

{kind=link}
{kind=link}
{kind=link}
羊肚菌胞外多糖快速估测方法
引用本文
陈莉, 孙永海, 付天宇, 丁健. 羊肚菌胞外多糖快速估测方法. 2014, 44(2): 567-572[CHEN Li, SUN Yong-hai, FU Tian-yu, DING Jian-feng. Method of rapid estimation of extracellular polysaccharide ofMorchella Esculenta . 吉林大学学报(工学版), 2014, 44(2): 567-572] 
Permissions
Copyright©2014, 吉林大学学报编辑部
版权所有.吉林大学学报编辑部
羊肚菌胞外多糖快速估测方法
陈莉(1983),女,博士研究生.研究方向:农产品智能检测与评价.E-mail:flygirl1983@126.com
摘要
针对羊肚菌深层发酵过程胞外多糖含量估测自动化关键技术问题,对发酵液的主要参数与胞外多糖相关性及其规律进行了分析,发现发酵液黏度、电导率与胞外多糖之间几乎不存在相关性。在0.01显著性水平下,发酵时间、发酵液浊度、残余还原糖与胞外多糖的相关性显著,相关系数均在0.7以上。在排除变量间的虚假相关后,只有发酵液浊度与胞外多糖的偏相关系数在0.8以上,可以认为发酵液浊度是描述胞外多糖的最佳参数。利用所获一元线性回归、二项式回归以及BP神经网络三种数学模型对胞外多糖含量进行的估测验证试验结果表明:BP神经网络模型的估测效果最好,估测方差仅为2.37×10-6。
关键词:
食品科学技术; 胞外多糖; 快速估测; 羊肚菌深层发酵; BP神经网络
中图分类号:TS207.3
文献标志码:A
文章编号:1671-5497(2014)2-567-6
Method of rapid estimation of extracellular polysaccharide ofMorchella Esculenta
Abstract
In order to rapidly estimate extracellular polysaccharide content in submerged fermentation process of Morchella Esculenta, relativities between the fermentation parameters and extracellular polysaccharide were analyzed. There is little relativity between viscidity or conductance in fermentation broth and extracellular polysaccharide. At 0.01 level of significance, the relationships of extracellular polysaccharide and fermentation time, turbidity in fermentation broth, residual reducing sugar are of perfect association that all the correlation coefficients are over 0.7. After false associations among the parameters are removed, only partial correlation coefficient of turbidity in fermentation broth and extracellular polysaccharide is above 0.8. So, turbidity in fermentation broth can been considered as the optimized characteristic parameter for extracellular polysaccharide. The three mathematic models by linear fitting, binomial regression, BP neural network were tested by estimation experiment of the extracellular polysaccharide content. The results show that the estimation by BP neural network is the best that the variance is only 2.37×10-6.
Keyword:
food science and technology; extracellular polysaccharide; rapid estimate; submerged fermentation ofMorchella Esculenta ; BP neural networks
0 引 言
羊肚菌多糖是羊肚菌重要的活性成分,具有降血脂、抗肿瘤等诸多功能[ 1]。许多学者对羊肚菌多糖的获取、分离、纯化等方法进行了有益探索,贾建会等[ 2]以深层发酵羊肚菌菌丝体和发酵液为主要原料提取多糖。欧超等[ 3]研究了羊肚菌发酵过程中碳源、氮源、无机盐、培养条件对菌丝体生物量、胞外多糖的影响以及发酵过程中菌丝体生物量等的动态变化。在羊肚菌深层发酵培养过程中,菌丝生物量和胞外多糖含量是两个最重要的追求目标,获得羊肚菌胞外多糖的传统方法是先将发酵液低温醇沉12 h,离心得胞外粗多糖,接着脱蛋白,再利用化学方法测得精确值,整个过程繁琐且耗时。如何实现在羊肚菌深层发酵过程中对菌丝生物量和胞外多糖含量两个指标的自动检测,对羊肚菌深层发酵生产质量控制有特殊意义。在对菌丝生物量自动检测方面,王莹等[ 4]利用图像处理技术,提出了基于BP神经网络的羊肚菌液体发酵量化方法,取得了有益的研究经验。而对羊肚菌胞外多糖的估测方法的研究多是理论探讨,武秋立等[ 5]研究了羊肚菌胞外多糖的发酵动力学模型,用Luedeking方程描述胞外多糖。王莹等[ 6]应用遗传算法快速对羊肚菌胞外多糖的动力学模型进行了参数估计,利用遗传算法对描述羊肚菌生长的Logistic模型和试验数据进行了拟合,目标函数为试验数据与模型的模拟数据的误差平方和,选用四阶龙格-库塔法解Logistic方程,遗传算法初始种群数为40,遗传代数为100代,遗传算法计算结果为:Logistic方程的最佳适应度为20,结果表明Logistic方程与试验数据拟合情况很好。这些方法虽然大大减少了估测时间,但需要测得相应时刻的菌体浓度,而试验过程中菌体浓度不太容易测得。本文通过对羊肚菌深层发酵培养过程中羊肚菌多糖与发酵液几个重要参数的相关性及其变化规律的分析,构建数学模型,以达到快速估测羊肚菌胞外多糖含量的目的。
1 材料与方法
1.1 材料
菌种:粗柄羊肚菌926号,购于四川绵阳食用菌研究所。
斜面培养基(PDA):称马铃薯200 g加水1000 mL煮沸30 min,四层纱布过滤,再配以葡萄糖20 g,3 g的KH2PO4,1.5 g的MgSO4,琼脂15 g,定容,混合均匀,pH值自然。
摇瓶发酵培养基:葡萄糖60 g/L,蛋白胨0.1 g/L,(NH4)2SO4为2 g/L,酵母膏5 g/L,KH2PO4为2 g/L,MgSO4为0.5 g/L,ZnSO4为0.1 g/L。初始pH值为5。
药品及试剂:0.1 mol/L的NaOH、0.1 mol/L的HCL DNS水杨酸溶液。95%乙醇,无水乙醇,蒽酮,浓硫酸。
仪器及设备:WGZ-100散射光浊度仪、PHS-3DPH计、数显黏度计、电导率仪、UV2300分光光度计、低速离心机LD4-A2。
1.2 试验方法
1.2.1 羊肚菌深层发酵工艺
(1)菌种活化:将4 ℃保存的粗柄羊肚菌母种接种于新鲜的PDA培养基上,置于25 ℃培养箱中培养48 h。
(2)摇瓶培养:选用1.1节中的摇瓶发酵培养基配方配好培养基,分装到50 mL的小三角瓶中,每瓶装料20 mL,用标准接种环向每个三角瓶中接入2块活化的约5 mm2大小的菌丝块,尽量保证菌块大小一致(所有的三角瓶近似看作平行样),然后置于28 ℃,100 r/min的空气浴振荡器上培养108 h。
1.2.2 羊肚菌发酵液参数的测定
按照1.2.1节方法做3轮重复试验,每轮培养108 h,间隔6 h取一个样,然后取每个时间点3个平行样的平均值作为最终的检测值。将每个发酵液样品以4000 r/min离心15 min,收集上清液,测量其浊度、黏度、电导率、残余还原糖和胞外多糖。随着发酵过程的进行,培养基的消耗程度有所变化,上述几个参数值也在变化。
发酵液的浊度利用WGZ-100散射光浊度仪测定。黏度利用数显黏度计测定。电导率利用电导率仪测定。
残余还原糖的测定方法:将发酵液离心后,用移液枪取上清液0.01 mL到25 mL比色管中,加入1.95 mL蒸馏水,再加入1.5 mL DNS水杨酸溶液,混和均匀;沸水加热5 min,取出,冷却至室温;补加21.5 mL蒸馏水,摇匀,静置20 min,利用分光光度计于540 nm处测吸光度。
胞外多糖的提取方法:将发酵液离心后,取上清液10 mL,加30 mL 95%乙醇于低温下醇沉12 h,再以4000 r/min离心15 min,弃去上清液,将沉淀干燥得粗多糖,其中含有蛋白质和多糖;然后用Savag法[ 7]将粗多糖脱蛋白,再进行低温醇沉12 h,冷冻干燥得纯多糖。
胞外多糖的测定方法:称取适量的胞外多糖纯品,加蒸馏水配成多糖溶液;取1 mL作为样品液滴于比色管中,将比色管放于冷水中,用滴定管沿壁慢慢加入0.2%的蒽酮试剂3 mL;沸水加热10 min;取出放于冷水中冷却10 min,再室温冷却10 min;利用分光光度计于540 nm处测吸光度。
2 胞外多糖相关性分析
2.1 羊肚菌生长曲线
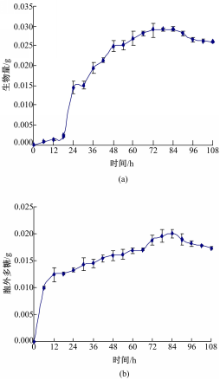
通过3次重复试验,每6 h取样,测定菌丝生物量和胞外多糖,对3次试验每个时间点的测量值取平均,做生长曲线,如图1所示:
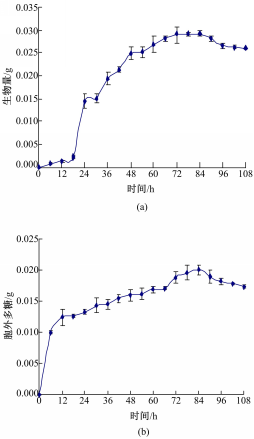 | 图1 羊肚菌生长曲线Fig.1 The growth curve of Morchella |
从图1(a)可以看出,0~12 h为菌体生长的缓慢期,菌丝量很少;12~72 h为菌体对数生长期,快速产生菌丝体;72~84 h菌体进入稳定期,产生大量菌丝体;84 h后进入衰亡期,菌丝量有所下降,产生自溶现象。从图1(b)可以看出,0~12 h胞外多糖产量很少;12~72 h快速积累了一些胞外多糖;72~84 h胞外多糖变化趋于平稳;84 h以后,胞外多糖产量呈下降趋势。胞外多糖属于羊肚菌发酵过程中的重要代谢产物,在它产生的过程中,必定会消耗培养基中的还原糖,还原糖的变化曲线如图2所示:
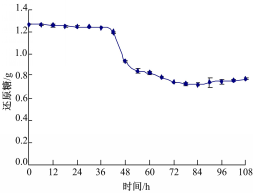 | 图2 还原糖变化曲线Fig.2 Change curve of reducing sugar |
从图2可以看出,在菌体的衰亡期之前,胞外多糖增加的过程中,还原糖一直处于下降趋势,衰亡期以后,胞外多糖开始下降。
2.2 发酵液参数与胞外多糖的相关性分析
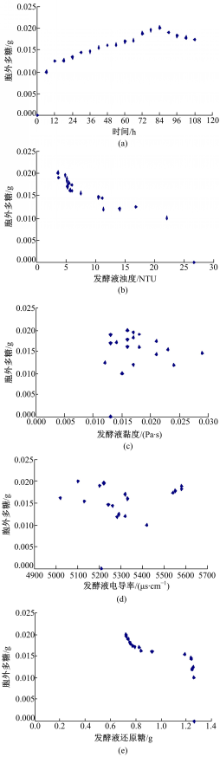
经过对预试验发酵过程的观察和分析,初选发酵时间、发酵液浊度、发酵液黏度、发酵液电导率以及残余还原糖作为描述胞外多糖含量的参数。为了分析这些参数与胞外多糖的相关性,每个时间点取3个平行样的均值为特征值,用SPSS17.0绘制散点图,如图3所示:
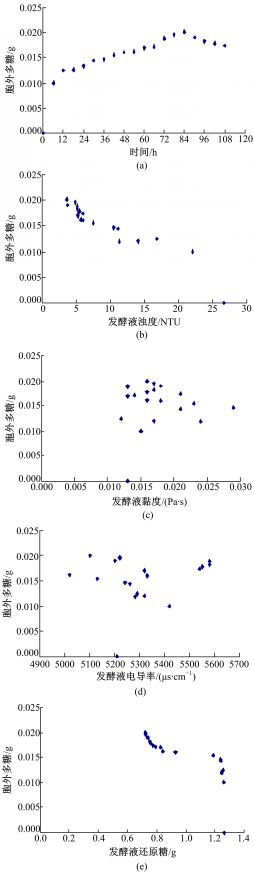 | 图3 胞外多糖与各参数关系散点图Fig.3 Scatter diagrams between characteristic value and extracellular polysaccharide |
| 表1 胞外多糖与各参数的相关系数 Table 1 Correlation coefficient between parameters and exopolysacharide |
表1给出了胞外多糖、发酵时间、发酵液浊度、发酵液黏度、发酵液电导率及残余还原糖各参数之间的相关系数。考虑到单纯利用相关系数来评价变量间的相关性还不够准确,还需要在剔除其他相关因素影响的条件下计算变量间的偏相关系数,如表2所示:
*表示在0.01水平(双侧)上显著相关。
| 表2 胞外多糖与各参数的偏相关系数 Table 2 Partial correlation coefficient between parameters and exopolysacharide |
由图3和表1中数据可知,发酵液的黏度、电导率与胞外多糖之间几乎不存在相关性。当显著性水平 a=0.01时,发酵时间、发酵液浊度、残余还原糖与胞外多糖的相关性显著,相关系数均在0.7以上。但在通过偏相关分析,排除变量间的虚假相关后,只有发酵液浊度与胞外多糖的偏相关系数在0.8以上(见表2),这可能是由于在羊肚菌生长过程中胞外多糖的形成消耗了发酵液中的营养物质,使得发酵液浊度随之变化所致,故最终选取发酵液浊度 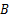
3 数学模型
根据2.2节发酵液各参数的相关性分析结果,选择发酵液浊度 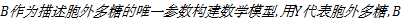
3.1 一元线性回归
在57组试验数据中取38组试验数据构造数学模型,利用SPSS17.0进行一元线性回归和二项式回归。
得到一元线性回归方程为
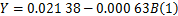
经检验得,复相关系数为0.945,判定系数 R2为0.894,回归方程估计标准误差为0.001 5311,常数和发酵液浊度的回归系数显著性检验的 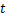
3.2 二项式回归
二项式回归模型为
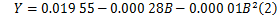
经检验得,回归方程显著性检验的 
4 模型估测效果对比
利用一元线性回归、二项式回归以及BP神经网络3种数据处理方法获得的数学模型对羊肚菌的胞外多糖含量估测效果进行试验验证。从试验数据中取19组数据,使用试验样本方差作为3种模型的评价指标来检验模型的准确性,检验结果如表3所示:
| 表3 三种模型检验结果 Table 3 Contrast results of three models |
样本方差计算公式为
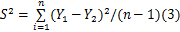
式中: 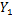
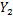
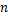
由表3可知,BP神经网络模型的检验方差值最低,即用BP神经网络模型预测胞外多糖的准确率最高,其次为二项式回归模型,一元线性回归模型的准确率最低。
5 结束语
通过对发酵时间、发酵液浊度、发酵液黏度、发酵液电导率、发酵液残余还原糖等参数与胞外多糖的相关性分析,发现发酵液的黏度、电导率与胞外多糖之间几乎不存在相关性。在0.01显著性水平下,发酵时间、发酵液浊度、残余还原糖与胞外多糖的相关性显著,相关系数均在0.7以上。但在排除变量间的虚假相关后,只有发酵液浊度与胞外多糖的偏相关系数在0.8以上,发酵液浊度 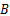
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|